Развитие насекомых. Эмбриональное развитие насекомых Эмбриогенез насекомых
Класс Насекомых отличается огромным разнообразием видов. Он подразделяется на два подкласса: подкласс Первичнобескрылых (Apterygota) и подкласс Крылатых насекомых (Pterygota).
В теле насекомых выделяются три отдела, или тагмы: голова, грудь и брюшко. Голова состоит из акрона и пяти слившихся сегментов; грудь образована тремя сегментами. В составе брюшка 10-12 сегментов и тельсон. Сегменты первых двух отделов имеют характерные придатки. В голове различают антенны, верхнюю губу, мандибулы, максиллы и нижнюю губу. Все сегменты груди имеют по одной паре ног. На дорсальной стороне второго и третьего сегмента груди крылатых насекомых образуются крылья. Сегменты брюшка обычно лишены конечностей.
По характеру развития крылатые насекомые подразделяются на две группы. Для представителей одних отрядов характерно постепенное превращение ювенильных постэмбриональных форм во взрослое животное. Для представителей других отрядов, напротив, характерно радикальное изменение личинки. Соответственно этому различают насекомых с неполным превращением (Hemimetabola) и с полным превращением (Holometabola). Согласно одной из гипотез, полное превращение является эволюционным приобретением и характеризует сравнительно молодые отряды. По-видимому, имеется определенная корреляция между характером развития и особенностями организации оогенеза. Если для Apterygota и Hemimetabola характерны паноистические яйцевые трубки, то Holometabola присущи в основном мероистические яичники, причем у перепончатокрылых, бабочек и мух это так называемые политрофные мероистические яичники. Такая корреляция примечательна. Она позволяет думать, что эволюционные преобразования, затрагивающие репродуктивную систему животных, важны не только непосредственно для репродуктивных процессов per se, но, открывая новые возможности для организации оогенеза, косвенным образом могут затрагивать сущностные события онтогенеза животных.
Деление ядра зиготы, которое окружено цитоплазматическим «двориком», происходит автономно, т. е. не сопровождается делением цитоплазмы. В результате синхронных кариокинетических делений возникают многочисленные ядра. Ядерные энергиды, т. е. ядра с окружающей их цитоплазмой, согласованно перемещаются по цитоплазматическим тяжам из глубоких слоев яйца на его периферию.
В период делений ядер и вплоть до их выхода на поверхность яйца зародыш имеет синцитиальную структуру. Это своеобразие ранних этапов развития имеет существенные физиологические последствия. Действительно, отсутствие разделяющих ядра клеточных мембран исключает возможность межклеточных взаимодействий с помощью лигандов и мембранных рецепторов, но обеспечивает свободную диффузию РНК и белков, запасенных в период оогенеза или синтезированных после оплодотворения, по всему зародышу, если они специально не зафиксированы в какой-либо области яйцевой клетки. Эти условия распространения сигнальных молекул у ранних зародышей насекомых резко отличаются от того, что имеется у животных, дробящиеся зародыши которых имеют клеточную структуру.
После выхода ядер в поверхностный слой цитоплазмы яйца - периплазму (у дрозофилы это происходит в конце 9-го цикла дробления), образуется синцитиальная бластодерма. В результате формирования мембран, разграничивающих клеточные территории, поверхность зародыша дробится на отдельные клетки («поверхностное» дробление) и возникает клеточная бластодерма.
Часть энергид дробления остается в желтке, становясь вителлофагами. Вителлофаги - это специализированные, часто полиплоидизированные клетки, которые участвуют в переработке и усвоении желтка. Образуемый вителлофагами желточный синцитий, возможно, играет и морфогенетическую роль, способствуя направленному разрастанию зародышевой полоски.
Зародышевая полоска представляет собой сгущение клеток бластодермы, которое обычно возникает на вентральной стороне яйца. Сгущение клеток происходит прежде всего благодаря изменению их формы - в области полоски они становятся столбчатыми. Различают три типа зародышевых полосок: короткие, полудлинные (промежуточные) и длинные. Короткие полоски имеют вид диска, материал которого дает начало сегментам головы, тогда как рост зародыша обеспечивается лежащей сзади от диска пролиферативной зоной. Полудлинные зародышевые полоски широко распространены у Hemimetabola, они образуют два вентролатеральных скопления клеток, которые смыкаются на вентральной стороне и дают начало голове и сегментам груди, тогда как остальные сегменты формируются за счет клеток зоны роста. Длинные полоски характерны для Holometabola. Они содержат материал всех сегментов и не имеют зоны роста, занимая не менее 80% длины яйца.
Как показывают наблюдения за судьбой клеток зародышевой полоски, ее медиальная часть на большем протяжении занята материалом презумптивной мезодермы, тогда как передняя и задняя области полоски - презумптивной энтодермой. По бокам от медиальных зачатков располагаются симметричные полоски нейроэктодермы.
Способы обособления зародышевых листков у насекомых многообразны. Погружение материала будущей мезодермы и энтодермы может происходить одновременно, но иногда эти процессы разобщены во времени. Разнообразны и механизмы перемещения клеточного материала с поверхности зародыша. У многих насекомых происходит иммиграция, т. е. выселение отдельных клеток, которые располагаются между эпибластом и желтком. У некоторых видов погружающиеся вглубь клетки не теряют эпителиальной структуры. Так, например, у Scolia quadripunctata имеет место своеобразная эпиболия. В ходе этого процесса медиальная часть зародышевой полоски отделяется продольными бороздами от ее латеральных частей. Последние своими свободными краями нарастают по поверхности отделившейся медиальной полоски навстречу друг другу и смыкаются, в конце концов, по средней линии зародыша. У многих насекомых, в том числе и у дрозофилы, происходит инвагинация медиальной части зародышевой полоски, в результате которой формируется продольно расположенная мезодермальная трубка. На стадии гаструляции в той области, где происходит иммиграция, инвагинация или эпиболия, некоторое время сохраняется продольная бороздка.
После отделения или инвагинации презумптивной мезодермы эктодерма латеральных областей зародыша смыкается по вентральной медиальной линии. Этот материал специфицирован как нейроэктодерма, которая служит источником нейронов и глиальных клеток и дает начало нервной системе насекомого. Нервные элементы иммигрируют с поверхности зародыша и располагаются под эктодермой. Нейроны каждого сегмента образуют ганглии, которые с помощью нейральных отростков объединяются в единую систему. Симметричные ганглии, лежащие в пределах одного сегмента, соединяются двумя комиссурами. С ипсилатеральными ганглиями, т. е. с ганглиями, лежащими на одной и той же стороне тела (от лат. ipse - сам), они соединяются с помощью коннектив.
Энтодермальные зачатки средней кишки образуются обычно на переднем и заднем концах зародышевой полоски. Впоследствии в области этих зачатков происходит инвагинация эктодермы и образуются - на переднем конце стомодеум, эктодермальный зачаток передней кишки, и на заднем конце проктодеум, эктодермальный зачаток задней кишки. На границе между задней и средней кишкой формируются органы выделительной системы насекомого - мальпигиевы трубочки, открывающиеся в заднюю кишку.
Одновременно с гаструляцией происходит растяжение зародышевой полоски. У видов с длинной зародышевой полоской в результате этого процесса она, распространяясь по срединной линии зародыша, огибает заднюю оконечность желточной массы и движется в переднем направлении по спинной стороне зародыша. При этом задний конец полоски приближается к ее переднему концу. В период растяжения полоска обнаруживает первые признаки метамеризации, поскольку образуются парасегменты и детерминируются границы будущих сегментов тела. На следующем этапе развития происходит укорочение зародышевой полоски, и ее задний конец вновь совпадает с задней оконечностью желточной массы.
Однослойная эктодерма, находящаяся за пределами зародышевой полоски, образует боковые складки, которые растут навстречу друг другу в вентральном направлении, смыкаясь по медиальной линии. В результате возникают две зародышевые оболочки. Внешний листок складки становится серозой (от лат. serosus — водянистый), которая распространяется на дорсальную область зародыша. Внутренний листок формирует амнион (от греч. - чаша, в которую собирали кровь жертвенного животного), эпителиальный пласт которого составляет единое целое с эктодермой зародыша. В результате этого процесса образуется замкнутая амниотическая полость, которая заполняется особой амниотической жидкостью, омывающей зародыш. Возникновение в процессе эволюции серозы и амниона обеспечило благоприятную среду для развития зародыша и, предотвратив опасность высыхания зародыша, послужило важным фактором завоевания суши насекомыми.
Предполагается, что формирование зародышевых оболочек первоначально возникло как следствие своеобразного перемещения зародыша в глубь яйца, процесса, который носит название бластокинеза. Бластокинез характерен для видов с короткой и полудлинной зародышевой полоской и состоит из двух фаз.
В первой фазе, которая обозначается как анатрепсис , происходит погружение зародыша в желток. Погружение начинается на заднем конце зародышевой полоски. Задний конец погружается внутрь желтка и поворачивается там на 180°. Благодаря движению задней области полоски в направлении переднего конца яйца зародыш приобретает сначала U-образную форму, а затем вновь принимает линейную форму, но теперь головная область погруженного в желток зародыша обращена назад. При этом перемещении связанная с зародышем внезародышевая эктодерма также увлекается в глубь желтка, в результате чего между вентральной стороной зародыша и желтком образуется амниотическая полость, ограниченная со стороны желтка однослойной внезародышевой оболочкой, амнионом. Остающаяся на поверхности яйца внезародышевая эктодерма представляет собой серозу.
Вторая фаза бластокинеза, или кататрепсис характеризуется обратным направлением процесса, в результате которого зародыш, теперь уже сегментированный и имеющий развитые придатки, выходит на поверхность. У эволюционно относительно молодых видов с длинной полоской истинный бластокинез отсутствует, хотя описанное выше растяжение и сокращение зародышевой полоски, в ходе которых также образуются амнион и сероза, видимо, можно рассматривать как процесс, имеющий общие корни с бластокинезом (О. М. Иванова-Казас, 1995).
В эмбриональный период происходит и формирование гонад. Если у представителей примитивных отрядов первичные половые клетки обособляются на сравнительно поздних стадиях развития, то у многих Holometabola, например у дрозофилы, они возникают, как уже говорилось, в период дробления и образования бластодермы.
Попадающие в заднюю область яйца ядра дробления окружаются полярной плазмой и обособляются в виде полярных клеток, которые как бы выталкиваются на поверхность яйца. В ходе гаструляции первичные половые клетки погружаются внутрь зародыша, локализуясь в заднем зачатке средней кишки. В период растяжения полоски они перемещаются на дорсальную сторону яйца и вместе с задней областью средней кишки смещаются вперед. Соматическая часть гонады формируется за счет латеральной мезодермы. В мезодермальном зачатке гонады экспрессируется ген columbus. Соответствующий белок служит аттрактантом, привлекающим мигрирующие первичные половые клетки. У дрозофилы зачаток гонады закладывается в V брюшном сегменте, при этом в состав гонады входит мезодерма V-VIII брюшных сегментов. Мезодермальные элементы проникают между гоноцитами и образуют фолликулярный эпителий яйцевых трубочек и сперматоцист.
Эмбриональный период у насекомых завершается появлением личинки. От взрослых насекомых вылупившиеся личинки отличаются прежде всего небольшими размерами и в тысячи раз меньшей массой. У личинок не полностью развита половая система, отсутствуют крылья и некоторые другие органы взрослых форм, или имаго (от лат. imago — изображение, портрет). Вместе с тем личинки имеют свои специфические органы, позволяющие им продолжать развитие, рост и накопление энергетических резервов, которые потребуются на стадии имаго для процессов размножения. Адаптивное значение личиночной стадии очевидно. Именно благодаря личиночной стадии становится реальным создание энергетического запаса, необходимого для достижения дефинитивного состояния и продолжения рода.
Поскольку покровный эпителий насекомых, как и у всех Ecdysozoa, вырабатывает жесткий кутикулярный экзоскелет, рост и развитие личинки возможны только благодаря периодическим линькам, в ходе которых покровы животного утрачивают связь с кутикулой. После линьки, или экдизиса (от греч. - удаление) открывается возможность роста. У насекомых, как и других членистоногих, имеются особые гормоны, контролирующие процессы линьки. Гормоном линьки служит экдизон, который вырабатывается в особых проторакальных (экдизиальных) железах личинок. Под влиянием экдизона в гиподерме личинки стимулируется пролиферация клеток и инициируется отделение кутикулы. Поддержание способности к линькам обеспечивается ювенильным гормоном, который продуцируется так называемыми прилежащими телами (corpora allata). Хирургическое удаление corpora allata ведет к преждевременному прекращению линек и формированию карликовых животных. Наоборот, трансплантация прилежащих тел от молодых личинок поздним продлевает период линек и способствует образованию ненормально крупных насекомых (Pflugfelder, 1958).
Начавшийся после линьки период роста завершается образованием экзоскелета, после чего рост останавливается. Периодическое сбрасывание кутикулярного покрова завершается по достижении имагинальной стадии. У некоторых насекомых число линек строго детерминировано и обычно невелико (до шести линек), у других число линек зависит от условий существования и достигает 20 (поденки) или даже 40 (платяная моль).
У Hemimetabola вышедшая из яйцевых оболочек молодь имеет значительное сходство с имаго, хотя и отличается от взрослых форм размерами и отсутствием крыльев, зачатки которых, впрочем, появляются уже после первой линьки. Ювенильные формы (от лат. juvenilis - юный) Hemimetabola называются нимфами (от греч. - девица, невеста) или (у веснянок, поденок, стрекоз) наядами (от греч. - нимфа вод). По мере развития нимфы растут и приобретают все характерные для имаго признаки.
У Holometabola личинки резко отличаются от взрослых форм. Как правило, они имеют червеобразное тело. У некоторых видов личинки лишены ног, у других имеют только грудные ноги, личинки третьих обладают и грудными и брюшными конечностями. В отличие от нимф личинки Holometabola в процессе роста и развития сохраняют свой личиночный облик. Трансформация в этом случае происходит на стадии куколки, когда возникают специфические признаки взрослого животного с характерным членением тела, с конечностями и крыльями. Появлению взрослого животного предшествует последняя линька.
В ходе развития многие ткани личинки полиплоидизируются и при превращении личинки во взрослую особь замещаются диплоидными. Глубина метаморфоза личинок варьирует. Например, у многих мух и перепончатокрылых в ходе метаморфоза практически вся личиночная гиподерма разрушается и замещается имагинальной. Разрушаются личиночные лабиальные железы. Радикальной перестройке подвергается мускулатура. Новый облик приобретает трахейная дыхательная система. Разрушение личиночных тканей происходит путем гистолиза и фагоцитоза. Активную роль в процессах метаморфоза играет апоптоз.
Построение новых органов взрослой особи происходит при участии специальных зачатков - так называемых имагинальных дисков, или особых резервных скоплений диплоидных имагинальных клеток - гистобластов, входящих в состав функционирующих полиплоидных тканей личинки. Дифференциация имагинальных дисков и гистобластов начинается при получении соответствующего гормонального сигнала на стадии куколки, тогда как спецификация клеток происходит на стадии бластодермы. Характер спецификации клеток, образующих впоследствии имагинальные диски, зависит от положения этих клеток вдоль передне-задней и дорсо-вентральной осей. Эти детерминированные на образование дисков клетки практически не пролиферируют, однако на личиночной стадии клеточные деления возобновляются, и соответственно происходит рост дисков. У дрозофилы имеется десять основных пар имагинальных дисков и один непарный диск в задней области тела, идущий на построение репродуктивных органов животного. Наиболее крупный диск дрозофилы - имагинальный диск крыла содержит около 60 тыс. клеток, имагинальные диски ног или жужжалец - примерно по 10 тыс. клеток.
Имагинальные диски закладываются у вылупившихся личинок в виде эпителиальных утолщений. Различают несколько типов дисков. Наиболее просто устроены наружный и свободный диски, располагающиеся непосредственно под кутикулой. Более сложно устроены погруженные диски. В этом случае гиподерма образует впячивание, погруженная часть которого образует собственно диск, соединенный с гиподермой периподиальным стебельком. На стадии куколки при действии гормона экдистперона происходит выворачивание имагинальных дисков наружу. При этом центральная часть диска телескопически выдвигается и образует дистальную область органа. Периферическая часть диска дает проксимальную область конечности.
Сразу после вылупления личинка нередко ещё лишена своей нормальной окраски, обычно бесцветная или беловатая и имеет мягкие покровы. Но у открыто живущих личинок окрашивание и затвердевание покровов происходят очень быстро, и личинка приобретает нормальный вид. Создаются стимулы питания, у некоторых - после переваривания остатков эмбрионального желтка и вывода экскрементов. Личинка вступает в пору усиленного питания, роста и развития. Рост и развитие сопровождаются периодическими линьками - сбрасыванием кутикулы . Благодаря линькам происходит увеличение тела и наружные его изменения.
Число линек в течение развития личинки изменяется от трёх (мухи) до 25-30 (подёнки и веснянки). После каждой линьки личинка переходит в следующий возраст , поэтому линьки разделяют между собой возраста личинок, при этом количество возрастов равно количеству линек.
Увеличение массы тела с каждой последующей линькой иногда подчиняется геометрической прогрессии со знаменателем q=2. Таким образом, масса тела личинки n-го возраста будет определяться следующей формулой:
A n = 2 n–1 a 1 ,
где a 1 - масса тела в первом возрасте. Однако такая зависимость наблюдается далеко не всегда: знаменатель прогрессии сильно зависит от условий питания и видовых особенностей. Так, например, покровы гусеницы сразу после линьки не прилегают плотно к телу, позволяя расти между линьками, и, например, за пять возрастов увеличивает массу тела в 10 000 раз.
Существенным является вопрос об определении возраста личинок. Без точного определения возрастов невозможно успешно организовать борьбу с вредителем, сигнализировать о сроках его развития или изучать биологию насекомого. Установление возраста по размерам тела ненадежно, так как в зависимости от состояния личинки её сегменты могут быть растянуты либо тесно сближены друг с другом, поэтому размеры личинок каждого возраста могут перекрываться соседними возрастами.
У насекомых с неполным превращением возраста личинок обычно хорошо различаются по ряду признаков - степени развития крыловых зачаток, числу члеников в усиках и др. Развитие личинок здесь сопровождается существенными внешними их изменениями.
У личинок насекомых с полным превращением переход их из одного возраста в другой проявляется главным образом в увеличении размеров тела, которые, однако, изменчивы. Поэтому для определения возраста у этих личинок обычно измеряют размеры твёрдых, нерастягивающихся частей тела, например, головной капсулы.
Типы метаморфоза у насекомых
Неполное превращение
Неполное превращение , или гемиметаморфоз (др.-греч. ἡμι- - «полу-» и μόρφωσις - «превращение»), в целом характеризуется прохождением лишь трёх стадий - яйца, личинки и имаго . Личинки насекомых с неполным превращением внешне сходны со взрослыми особями и, подобно последним, имеют сложные глаза, такие же, как у взрослых, ротовые органы и в более старших возрастах - хорошо выраженные наружные зачатки крыльев . Помимо этого, у многих насекомых с неполным превращением личинки ведут сходный с имаго образ жизни и могут встречаться совместно с последними. Насекомых с этим типом метаморфоза относят к таксономической группе Exopterygota (англ. ) (или Hemimetabola ) в составе инфракласса Новокрылых .
Вследствие большого морфологического и биологического сходства с имаго такие личинки называются имагообразными или нимфами (встречающееся иногда применение термина «нимфа» к личинкам лишь последних возрастов с развитыми крыловыми зачатками является некорректным). Нимфы веснянок и стрекоз претерпевают особую модификацию - они также похожи на имаго, но живут в воде и обладают провизорными , то есть специально личиночными, органами - жабрами и др. Таких личинок называют наядами .
В качестве подтипа гемиметаморфоза выделяют также его упрощение - гипоморфоз (лат. hypomorphosis ), характерный для вторично бескрылых представителей насекомых с неполным превращением - вшей , пухоедов , некоторых тараканов , прямокрылых и др. При гипоморфозе нимфы внешне практически неотличимы от имаго. Прежде считалось даже, что эти насекомые развиваются без превращения.
Полное превращение
Полное превращение , или голометаморфоз (др.-греч. ὅλος - целый, полный и μεταμόρφωσις - превращение), характеризуется прохождением от четырёх до пяти стадий - яйца, личинки, куколки , имаго и иногда предкуколки . Насекомых с этим типом метаморфоза относят к таксономической группе Endopterygota (или Holometabola ) в составе инфракласса Новокрылых .
Личинки насекомых с полным превращением, как правило, мало похожи на взрослых особей, поэтому называются истинными личинками . Действительно, трудно узнать в червеобразной личинке жука или гусенице бабочки взрослое крылатое насекомое. При этом личинка всегда лишена сложных глаз, наружных зачатков крыльев и часто имеет иной тип ротовых органов, нежели взрослые особи. Нередко сегменты тела истинной личинки однородны, то есть отсутствует резкое разделение тела на грудной и брюшной отделы. Помимо того, истинные личинки живут в иной среде, чем взрослые. В связи с этим большинство органов истинных личинок имеют временный или, как говорят, провизорный характер, выполняя функции чисто личиночной жизни. От этих органов у взрослых насекомых обычно не остается и следа. К числу провизорных органов личинок относятся: брюшные ноги, ротовой аппарат, паутинные железы и т. д.
Основная масса истинных личинок относится к трем группам:
- Камподеовидные - внешне сходны с двухвостками , подвижные, обычно темноокрашенные личинки с плотными покровами и тремя парами грудных ног, хорошо обособленной прогнатической головой и часто со щетинками на конце тела (жужелицы , плавунцы , сетчатокрылые и др.);
- Червеобразные - малоподвижные, светлоокрашенные личинки, лишенные брюшных и грудных ног или с короткими грудными ногами (двукрылые , многие перепончатокрылые , многие жуки);
- Гусеницеобразные , или эруковидные , - умеренно подвижные личинки с тремя парами грудных ног и 2-8 парами брюшных (личинки скорпионниц , гусеницы , ложногусеницы).
Согласно теории Берлезе, эмбрион проходит в яйце несколько стадий, на которых возможно вылупление. Основанная на этой концепции классификация делит личинок по этим стадиям.
Несходство истинных личинок со взрослой фазой распространяется и на образ жизни - питание, место обитания и пр.
Закончив свой рост, истинная личинка последнего возраста прекращает питание, становится неподвижной, линяет в последний раз и превращается в куколку . Иногда куколке предшествует особая стадия, называемая предкуколкой. По сути это последний возраст истинной личинки, переход в который сопровождается утратой подвижности, сокращением размеров и началом процесса гистолиза задолго до линьки в куколку.
Анаморфоз
Физиология метаморфоза
Метаморфоз сопровождается, помимо внешних, также и внутренними изменениями. Важнейшую роль при метаморфозе играют гормоны .
При неполном превращении внутренние изменения протекают постепенно и при переходе во взрослую фазу не сопровождаются коренной перестройкой всей личиночной организации; многие органы личинок сохраняются у имаго. Даже зачатки яичников и семенников обнаруживаются уже у личинок первого возраста, развиваются постепенно и при переходе во взрослую стадию у одних видов превращаются во вполне зрелые половые железы, у других доразвиваются во взрослом состоянии.
Совсем иной характер внутренних изменений имеют насекомые с полным превращением. Их личинки резко отличаются от имаго, поэтому переход во взрослую стадию неизбежно требует коренной перестройки всей морфологической и биологической организации. Эта перестройка происходит в основном в стадии куколки и слагается из двух процессов - гистолиза и гистогенеза .
Сущность гистолиза состоит в уничтожении личиночных органов, сопровождаемым проникновением и внедрением в ткани гемоцитов. Содержимое куколки как бы возвращается к недифференцированному состоянию яйца. Гистолизу подвергаются все системы организма личинки, кроме нервной, половой, а также спинного сосуда.
В дальнейшем гистолиз сменяется гистогенезом, конечной целью которого является построение из образовавшейся жидкой массы новых, имагинальных органов. Важную роль при гистогенезе также играют имагинальные зачатки - группы клеток, из которых возникают те или иные ткани и органы.
Существеннейшую роль при метаморфозе играет эндокринная система . Рост и развитие насекомых регулируются нейросекреторными клетками головного мозга, кардиальными телами, прилежащими телами и переднегрудными железами. Мозговой гормон через аксоны мозга переносится к кардиальным телам, а оттуда - в кровь . Он стимулирует обмен веществ , а также переднегрудные железы, вырабатывающие гормон линьки экдизон . Прилежащие тела выделяют ювенильный гормон , который препятствует линьке во взрослую фазу и стимулирует рост и развитие личиночных органов. С возрастом рост тела личинки опережает рост ювенильных тел, и их роль постепенно затухает. Поступление ювенильного гормона в кровь прекращается и насекомое вступает в стадию имаго.
Результатом проводившихся экспериментов по пересадке ювенильных тел в личинку последнего возраста у прямокрылообразных и клопов стало получение дополнительных, ненормально крупных личинок. Так в условиях опыта удается увеличить число личиночных возрастов.
Фазовая изменчивость
Особую форму метаморфоза, проходимую совместно с основным, представляет собой так называемая фазовая изменчивость , открытая первоначально у саранчовых , но затем обнаруженная и у остальных прямокрылых , а также у палочников , бабочек и других насекомых. Сущность её заключается в том, что при возникновении скученности особей изменяются морфологические и биолого-физиологические свойства вида - усиливается пигментация покровов (в основном из-за частичного отложения в них продуктов метаболизма), изменяются пропорции и форма частей тела, ускоряется развитие, меняется поведение. Возникает стадная фаза (лат. phasis gregaria ). При рассеивании особей эти свойства утрачиваются и вид возвращается через переходную фазу (лат. phasis transiens ) в свое исходное состояние - одиночную фазу (лат. phasis solitaria ). Отличия между фазами столь велики, что незнание их природы порождало разделение одного вида на 2-3.
Происхождение полного превращения
Несомненно, что полное превращение явилось гигантским скачком в эволюции насекомых и всего типа членистоногих . Относительно его возникновения существует две основных гипотезы. Сторонники обеих согласны, что неполное превращение и, соответственно, его очевидные производные (гипо- и гиперморфоз) произошли от первичного типа метаморфоза - протоморфоза. Что касается возникновения голометаморфоза, мнения расходятся.
Согласно гипотезе независимого происхождения , предложенной М. С. Гиляровым , А. А. Захваткиным и А. Г. Шаровым , полное превращение является эволюционным развитием протоморфоза. При протоморфозе личинки и имаго обычно живут в почве, под растительными останками и в других защищённых местах. Но приобретение крыльев неизбежно способствовало переходу имаго к открытому образу жизни, что, в свою очередь, вынуждало их нежных личинок также жить открыто, под сильным воздействием неблагоприятных условий. Возникла необходимость приспособления личинок к этим условиям, которое пошло в итоге двумя путями. В одних случаях развитие стало всё более проходить под защитой яйца. Вылупление всё более запаздывало и происходило на более поздних стадиях развития, то есть осуществлялась эмбрионизация развития. Личинки вылуплялись уже приспособленными к открытому образу жизни, само же яйцо стало намного крупнее и богаче питательным желтком. В других случаях личинки перешли к ещё более скрытому образу жизни, благодаря чему у них всё более усиливались червеобразные признаки, происходила деэмбрионизация зародыша. Возник резкий разрыв между стадией личинки и имаго, который был восполнен стадией куколки. Так возник голометаморфоз.
Сторонники второй гипотезы предполагают, что полное превращение развилось из неполного. Эти представления связаны, в первую очередь, с именами итальянского учёного А. Берлезе и русского исследователя И. И. Ёжикова. По их мнению, личинки насекомых с полным превращением возникли вследствие выхода из яйца на более ранних стадиях развития. Образно говоря, истинные личинки являются свободно живущими эмбрионами . Отличие от предыдущей теории в том, что стадия куколки не является новой стадией в полном смысле слова, а соответствует совокупности нимфальных стадий насекомых с неполным превращением. Одной из причин эмбрионизации считается малое содержание желтка в яйцах насекомых с полным превращением, в результате чего личинка не успевает завершить своё развитие в яйце и вынуждена вылупляться раньше. Противоположно, яйца насекомых с неполным превращением, как более крупные, содержат больше питательного желтка. Поэтому зародыш в своём развитии достигает более поздних стадий и вылупляется уже с признаками имаго.
Существует и третья, менее распространенная гипотеза, согласно которой истинные личинки являются прямыми модификациями нимф, то есть что голометаморфоз развился из гиперморфоза. Причину такой модификации видят в дифференцировании среды обитания и образа жизни личинок и имаго.
Некоторые положения этих гипотез подкрепляют друг друга, другие являются взаимоисключающими. Всё это говорит о недостаточности наших знаний об эволюции метаморфоза.
Функциональная роль стадий метаморфоза
Для насекомых характерно разделение функций не только в пространстве (дифференциация органов), но и во времени: в течение онтогенеза происходит не только смена функций отдельных органов, но меняются и основные функции организма как целого.
Для личиночных стадий насекомых характерна функция питания, накопления пластических и энергетических веществ, а для имаго - функция размножения. Функция расселения может выполняться как имагинальной, так и личиночными стадиями. Большинству насекомых, как несомненно прогрессивной группе, свойственно активное расселение, осуществляемое полётом. Поэтому весь облик имаго большей части крылатых насекомых - это облик расселяющейся стадии.
Иногда имагинальная стадия выполняет только функции видовой жизни - расселительную и размножения , утрачивая функцию питания. Однако выполнение функций расселения и размножения часто протекает в различные сроки; часто расселение происходит в те сроки, когда половые продукты ещё не созрели. Так, самки многих совок совершают перелёты, когда их яичники ещё не созрели. Более того, после созревания половых продуктов они отяжелевают настолько, что теряют способность даже к небольшим взлётам. Расселительную функцию иногда имеют и брачные полёты, особенно у общественных насекомых . Таким образом, у многих насекомых можно говорить о возрастном функциональном разделении стадии имаго на две «подстадии»: расселительную и репродуктивную.
Развитие насекомых делится на два периода: зародышевое, или эмбриональное (внутри яйца), и внеяйцевое, или постэмбриональное. Развитие зародыша в яйце начинается дроблением ядра с момента оплодотворения. Яйцо имеет круглую, овальную, и др. форму и представляет собой одну крупную клетку, включающую питательный желток, необходимый для роста и развития зародыша. Снаружи яйцо покрыто оболочкой (хорион), играющей роль скорлупы. На одном из полюсов яйца имеется одно или несколько мельчайших отверстий (микропиле), через которые проникают сперматозоиды при оплодотворении. Яйцо после откладки часто всасывает влагу из окружающей среды и вследствие этого увеличивается в 2-3 раза.
Зрелые яйца самки откладывают поодиночке или группами на листья, ветви и стволы деревьев, на почву, травянистую растительность и другие предметы. Часто яйца бывают погружены в субстрат (в древесную ткань или почву) или защищены пушком, снятым с конца -брюшка самки (у златогузки, непарного шелкопряда и других бабочек), или щитком, образовавшимся из выделений придаточных желез (у зеленой узкотелой златки).
Развитие зародыша в яйце продолжается от нескольких дней до месяца и более. Оно заключается в ряде сложных преобразований и заканчивается вылуплением личинки, прогрызающей оболочку яйца
Строение яйца
Яйцо насекомого представляет собой крупную клетку, покрытую снаружи хорионом, обычно имеющим характер скорлупы, под которой находится истинная, или желточная, оболочка. Хорион образован из выделений эпителиальных клеток яйцевой трубки яичника, и на его поверхности часто можно обнаружить более или менее сильно выраженные скульптурные образования, представляющие отпечатки клеток фолликулярного эпителия. На поверхности хориона имеется отверстие - микропиле, которое служит для проникновения сперматозоида.
Типы яиц и характер их откладки. Кроме размера, яйца могут различаться формой, скульптурой хориона, окраской. По форме они чаще бывают овальными, удлиненно-овальными, шаровидными. Вместе с тем в ряде случаев форма яиц очень типична для нескольких видов одного рода, а нередко и семейства. Так, очень характерны бутылковидные яйца у капустной и репной белянок, бочонковидные - у клопов щитников, полу шаровидные - у совок и т. д.
Наружная поверхность яиц, т.е. скульптура хориона, чаще гладкая, но может иметь продольные или поперечные ребрышки, морщинки, бороздки и т. д. Так, у яиц бабочек совок хорион имеет радиальные ребрышки, у яиц капустной и репной белянок - крупные продольные и мелкие поперечные ребрышки, у яиц некоторых кокцинеллид - мелкоячеистую скульптуру и т. д.
Самки насекомых откладывают большое количество яиц в один или несколько приемов. При этом размещают их по одному, небольшими группами или кладками по 100, 200 и более яиц, что типично для данного вида. Так, капустная белянка откладывает яйца на листья растений кладками до 200 яиц в каждой, а репная белянка - по одному.
Кладки яиц могут быть отложены открыто на растениях, например у капустной белянки, кольчатого шелкопряда, кокцинеллид, или скрыты в субстрате (почва, ткани растений), прикрыты волосками с брюшка самки или снаружи защищены стенками кожистой капсулы. Так, кладки яиц непарного шелкопряда и златогузки настолько плотно прикрыты волосками с брюшка самки, что сверху совершенно не видны, а кладки яиц тараканов и богомолов находятся внутри кожистой капсулы - оотеки, образованной застывшими выделениями придаточных желез самки. Очень своеобразны также кубышки саранчовых, содержащие от 5-15 яиц у атбасарки до 140 яиц у пустынной саранчи. Секрет придаточных желез самки не только обволакивает их, но и цементирует прилегающие частицы почвы.
Развитие зародыша
Эмбриональное развитие начинается с дробления ядра и передвижения образовавшихся дочерних ядер с небольшими участками протоплазмы к периферии яйца. Здесь они образуют сплошной слой клеток - бластодерму, охватывающую скопление желтка. В дальнейшем бластодерма дифференцируется на зародышевую и внезародышевую зоны. Клетки зародышевой зоны делятся более интенсивно и образуют с брюшной стороны яйца зародышевую полоску.
В процессе дальнейшей дифференциации зародышевой полоски образуется ее наружный листок - эктодерма, по краям формирующая двухслойную складку, а сама полоска несколько погружается в желток. Нарастающие края складки смыкаются друг с другом и срастаются, в результате чего зародышевая полоска покрывается двумя слоями клеток - зародышевыми оболочками. Наружную оболочку называют серозой, внутреннюю - амнионом. Между амнионом и развивающимся зародышем образуется полость, заполненная жидкостью, имеющей защитное и пищевое значение для зародыша. Наличие оболочек объясняет повышенную устойчивость яиц насекомых после прохождения этого этапа эмбриогенеза к инсектицидам, а также к иссушению.
По мере образования зародышевых листков начинает формироваться зародыш насекомого. Из эктодермы образуются кожные покровы тела; путем впячивания эктодермы с обоих полюсов возникают ротовое и анальное отверстия, передний и задний отделы кишечника, зачатки будущих трахей и нервной системы. Из внутреннего зародышевого листка - энтодермы образуется эпителий средней кишки, из среднего - мезодермы формируются мышцы, жировое тело, спинной сосуд, оболочка половых желез.
Сформировавшийся зародыш, превратившийся, по существу, в личинку, заполняет все яйцо, начинает интенсивно двигаться, набирает воздух в трахеи, заглатывает амниотическую жидкость, увеличиваясь в объеме, освобождается от зародышевых оболочек, разрывает хорион и вылупляется, т. е. выходит наружу.
Обычно эмбриональное развитие насекомых длится от 2 до 10 дней, реже 2-3 недели, в зависимости главным образом от температуры среды. При длительной остановке в развитии - эмбриональной диапаузе - продолжительность фазы яйца у некоторых насекомых (саранчовые, мухи цветочницы и др.) может достигать 6-9 месяцев.
Рис. 1. Развитие насекомого с неполным превращениемУ насекомых существует два основных типа развития, в соответствии с которыми их подразделяют на два крупных отдела.
Развитие с неполным превращением проходит в три стадии: яйцо - личинка - имаго (взрослое насекомое), не сопровождаясь резкими изменениями (рис. 1 ). Личинки по строению подобны взрослым насекомым и превращаются в них сразу после окончания развития и последней линьки.
Развитие с полным превращением проходит в четыре стадии: яйцо - личинка - куколка - имаго. Личинки резко отличаются от имаго, поэтому, окончив развитие, они превращаются в покоящуюся стадию куколки, в которой происходит преобразование организации тела от личиночной до имагинальной.
Строение и формы яиц
Яйца насекомых богаты желтком и покрыты двумя оболочками. Они разнообразны по размерам, форме и окраске. Размеры яиц обычно составляют от десятых долей миллиметра до нескольких миллиметров. По форме яйца бывают овальными (многие жуки, бабочки, мухи), удлиненными (кузнечики и саранчовые), шаровидными (клопы-черепашки), полушаровидными (бабочки-совки), бочонковидными (клопы-щитники), бутылковидными (дневные бабочки), стебельчатыми (златоглазки) (рис. 2 ).
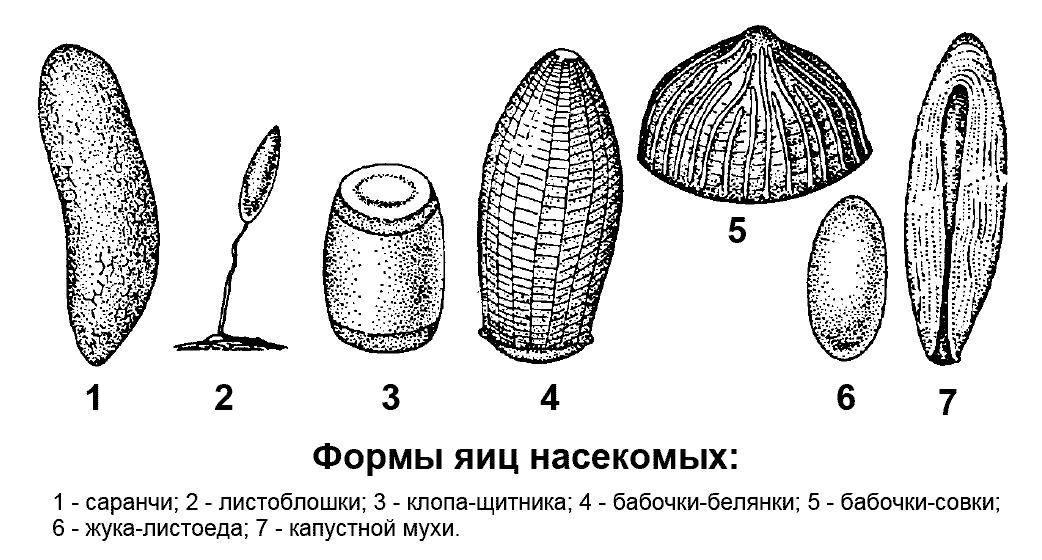 Рис. 2. Формы яиц насекомых
Рис. 2. Формы яиц насекомых Самки различных насекомых откладывают яйца в места, пригодные для дальнейшего развития личинок. Плодовитость насекомых (количество яиц, откладываемое самкой в течение жизни) разнообразна. У некоторых видов она невелика - порядка нескольких десятков яиц, у большинства около нескольких сотен, у ряда видов - несколько тысяч. Плодовитость половозрелых самок общественных насекомых достигает нескольких миллионов яиц. У некоторых насекомых самки откладывают яйца одиночно или врассыпную, однако большинство откладывает их компактными группами - яйцекладками.
Строение и типы личинок
Основные функции личиночной стадии - питание, рост, развитие. В развитии насекомых личинка - единственная растущая стадия. Ее размеры могут увеличиваться в десятки и сотни раз. Поэтому личинки несколько раз проходят линьку, сбрасывая старую кутикулу в виде личиночной шкурки и одновременно образуя новую, рассчитанную на дальнейший рост.
Имагообразные линийки, или нимфы, - личинки насекомых с неполным превращением. Эти личинки сходны по строению со взрослыми насекомыми. Они имеют на грудном отделе три пары хорошо развитых ног, фасеточные глаза, ротовой аппарат, подобный аппарату имаго, хорошо развитые антенны. У них развиваются наружные зачатки крыльев. Обычно имагообразные личинки сходны с имаго также по образу жизни и характеру питания. У вредителей с неполным превращением личинки и имаго всегда повреждают растения сходным образом. Некоторое исключение составляют водноживущие личинки стрекоз и поденок, имеющие органы водного дыхания - трахейные жабры. Их иногда называют наядами.
Личинок насекомых с полным превращением иногда называют истинными . Эти личинки могут иметь различное количество конечностей, развитых слабее, чем у взрослых насекомых; либо конечности отсутствуют. Личинки лишены фасеточных глаз, органами зрения служат простые глазки. У них отсутствуют наружные зачатки крыльев, антенны короткие или неразвиты, ротовой аппарат чаще всего грызущий, он может быть совершенно иного строения, нежели у имаго. Личинки насекомых с полным превращением могут резко отличаться от имаго по образу жизни и характеру питания. У вредителей с полным превращением вред может наносить либо личинка, либо имаго, или насекомые в обеих стадиях развития.
У насекомых с полным превращением выделяют три типа личинок (рис. 3 ).
 Рис. 3. Типы личинок насекомых с полным превращением
Рис. 3. Типы личинок насекомых с полным превращением Камподеовидные личинки (от лат. названия первичнобескрылого насекомого камподея, сходного с представителями данного типа) - активные, нередко хищные, с развитыми придатками тела личинки. У них хорошо развиты три пары грудных ног, полностью расчлененных, длина их обычно превышает ширину тела. Сравнительно четко выделены грудной и брюшной отделы. Развиты антенны, характерны крупные верхние челюсти. Покровы обычно пигментированы. Этот тип личинок встречается у некоторых семейств отряда Жесткокрылые (жужелицы, стафилины, божьи коровки) и у всех Сетчатокрылых.
Червеобразные личинки - очень крупная и неоднородная группа. Тело у них удлиненное, слабо подразделено на грудной и брюшной отделы. Три пары грудных ног короткие, либо отсутствуют. По наличию ног и развитию головы выделяют три формы червеобразных личинок. Личинки с головой и ногами встречаются у насекомых ряда семейств отряда Жесткокрылые и имеют разнообразную внешность. Личинки Пластинчатоусых (жуки-хрущи и навозники) толстые, С-образно изогнуты; личинки жуков-листоедов заметно выпуклые сверху; личинки жуков-щелкунов и чернотелок (проволочники и ложнопроволочники) длинные, тонкие, цилиндрические, с уплотненными покровами. Личинки с головой и без ног встречаются также у некоторых представителей семейств отряда Жесткокрылые (златки, усачи, долгоносики), у большинства групп отряда Перепончатокрылые (наездники, осы, пчелы, муравьи) и у части видов отряда Двукрылые (большинство комаров). Личинки без ног и внешне выраженной головы есть только у ряда представителей в отряде Двукрылые (большинство мух). Головной отдел этих личинок втянут в переднегрудь, головная капсула недоразвита.
Гусеницеобразные личинки имеют удлиненное тело и большое количество коротких ног. В отличие от других личинок, у них помимо трех пар грудных ног развито несколько пар придаточных брюшных ног. Эти короткие нерасчлененные личиночные придатки у взрослых насекомых не проявляются. Выделяют две формы гусеницеобразных личинок. Настоящие гусеницы - личинки Чешуекрылых имеют от 2 до 5 пар брюшных ног. Гусеницы большинства бабочек имеют 5 пар ног, расположенных на 3 -6-м и 9-м сегментах брюшка. Однако у некоторых семейств число брюшных ног сокращается до 4 (хохлатки), 3 (совки-металловидки) и 2 пар (пяденицы). Ложно гусеницы - личинки некоторых семейств пилильщиков, представителей отряда Перепончатокрылые. У них развито большее количество брюшных ног -6-8 пар.
Строение и типы куколок
У насекомых с полным превращением личинка, окончив развитие, превращается в куколку. Личинки большинства насекомых окукливаются в почве, иногда открыто - на растениях или внутри них. Куколка - покоящаяся стадия, однако в ее теле идут интенсивные противоположные процессы: распад личиночных органов и тканей и формирование органов и тканей имаго. К концу развития куколка приобретает основные черты строения взрослого насекомого: развитые крылья, ноги, антенны, ротовой аппарат имаго, фасеточные глаза. Придатки компактно сложены и прижаты к телу. Тело нередко покрыто дополнительной защитной оболочкой. По этому признаку выделяют три основные типа куколок (рис. 4 ).
 Рис. 4. Типы куколок насекомых
Рис. 4. Типы куколок насекомых Открытые , или свободные, куколки не имеют оболочки; все придатки тела хорошо различимы, свободны. Наглядно проявляется сходство куколок с имаго. Этот тип куколок встречается у подавляющего большинства Жесткокрылых и Перепончатокрылых, а также у насекомых отряда Сетчатокрылые. Очень редко открытые куколки встречаются у Чешуекрылых и Двукрылых.
Покрытые куколки имеют тонкую кожистую полупрозрачную оболочку, через которую можно различить ноги, крылья, антенны и другие части тела. Оболочка образуется выделениями кожных желез личинки в момент окукливания. Этот тип куколок характерен для подавляющего большинства Чешуекрылых, части отряда Двукрылые, очень редко встречается у Жесткокрылых и Перепончатокрылых.
Скрытые куколки заключены в плотную непрозрачную оболочку - пупарий, через которую нельзя различить части тела. Эта оболочка представляет собой личиночную шкурку, которая не сбрасывается как обычно при окукливании, а только отслаивается от тела и служит защитным покровом. Скрытая куколка характерна для большей части Двукрылых (большинство мух).
Эмбриональное развитие насекомых в типичном случае начинается с поверхностного дробления яйца, что связано с богатым содержанием у последнего питательного желтка. Образовавшаяся на поверхности яйца бастодерма на некотором участке поверхности толще; это утолщение дает зародышевую полоску и определяет брюшную сторону зародыша.
Образующаяся вдоль края зародышевой полоски кольцевая складка, обрастая и срастаясь над зародышевой полоской, дает два слоя зародышевых оболочек — внутреннюю (амнион) и наружную (серозу). От пласта клеток зародышевой полоски на стороне, обращенной к желтку, обособляется первичная энтодерма — общий зачаток энтодермы и мезодермы. В дальнейшем зародышевая полоска сегментируется.
Постэмбриональное развитие насекомых весьма разнообразно в связи с разнообразием условий существования молоди насекомого и степени ее приспособления к ним. Можно различать два основных типа постэмбрионального развития: неполное превращение—гемиметаболию и полное превращение — голометаболию. Голометаболия характеризуется глубокими изменениями молоди, в результате которых молодь по своему строению резко отлична от взрослого насекомого (имаго). Поэтому переход от состояния молоди, т. е. от личиночной фазы к фазе имагиналь-ной, требует коренной перестройки органов; это достигается при участии промежуточной фазы между фазами личиночной и имагинальной, в течение которой молодь сохраняет покой; эта промежуточная фаза — куколочная. В течение куколочного состояния внутренние органы насекомого либо подвергаются большей или меньшей перестройке, либо подвергаются полному разрушению (гистолизу) и строятся заново; в последнем случае образование органа заново происходит за счет так называемых имагинальных дисков, состоящих из клеток эмбрионального характера (гистобластов). Таким образом, куколочный период — период деятельного гистолиза и гистогенеза (новообразования тканей). Внешне куколка представляет имагинальную фазу с характерными для нее органами, которые, однако, остались недоразвитыми.
Голометаболично развитие в высших отрядах насекомых у жуков, чешуекрылых, ручейников, блох, перепончатокрылых и двукрылых.
В случае неполного превращения — гемиметаболии — столь глубокого различия в строении молодой и взрослой фазы нет: молодь то более, то несколько менее напоминает по строению имагинальную фазу Наконец, в тех случаях, когда переход от молодой фазы к взрослой сводится лишь к росту и сбрасыванию (в связи с ростом) старого покрова (экзувия), говорят об аметаболии, т е. отсутствии превращения; примером аметаболии служит развитие Collembola среди Apterygota. Однако обычно с последовательными возрастами, которые отделены друг от друга линьками, т е. сбрасыванием покрова предшествующего возраста, происходят и изменения в форме органов: так, например, относительно увеличивается длина усиков и церок, увеличивается число члеников; могут и вновь появляться некоторые органы, как, например, наружные половые придатки. Таково развитие у некоторых Apterygota, как-то: Thysanura и Diplura и у большинства гемиметаболичных Pterygota. У некоторых Pterygota, развивающихся с гемиметаболией (стрекоз, поденок; веснянок и части членистохоботных), развитие осложняется в связи с тем, что молодь живет в иных условиях, чем имаго; так, молодь стрекоз, поденок и веснянок развивается в водоемах и в связи с этим приобретает органы водного дыхания — жабры, которые представляют пример личиночных (провизорных) органов. Условия существования отражаются и на развитии других органов личинки, которые приспосабливаются к обстановке, например превращение в копательный орган передних ножек молоди цикады в связи с ее подземной жизнью.